Altruism and Cheating in Biofilms
Altrusim
Altruism is often described as a selfless concern for the welfare of others. It includes actions taken by one individual for the exclusive benefit of another even at the expense of the altruist. Jan-Ulrich Kreft, 2004 defines altruism as “behavior that increases the fitness of the group relative to other groups while it decreases the fitness of the altruist relative to others within the group.” In human society altruism is often considered a basic social virtue. Some, however, have questioned if altruism really exists in humans or for that matter in any other species.
Traditionally, altruism in human culture is explained either on the basis of reciprocity or kin selection. Reciprocity suggests that “altruistic acts” performed by one member of a culture are done consciously or unconsciously in expectation of a reciprocal “altruistic act” at some point in the future. One might ask, if there is truly an expectation of a reciprocal “altruistic act” can this really be considered altruism?
In kin selection it is hypothesized that an individual will be willing to put themselves at risk in proportion to the degree of relatedness of the beneficiaries of their intervention. In an evolutionary sense, sacrificing your life in order to save two brothers or eight cousins makes evolutionary sense since those individuals carry as many of the same genes into future generations as you do yourself. Among single celled organisms we have no reason to expect sentience or the ability to anticipate future acts of “kindness” on the part of other microorganisms, therefore we should probably seek any hint of altruism among hypotheses related to kin selection.
Examples of altruistic behavior have been proposed for a diverse variety of microorganisms including algae (Volvox), the cellular slime molds (Dictyostelium) and the eubacteria (Bacillus, Myxococcus, Pseudomonas). If, as suggested, this array of organisms does in fact display characteristics that can be considered altruistic, it suggests that this sort of cooperative behavior arose very early in the evolution of life on earth.
As described above (Multicellularity) Myxococcus xanthus is a motile bacterium with an intriguing collection of abilities. The organism swarms while “hunting” for its bacterial prey. It releases antibiotics and bacteriolytic compounds, which kill and digest these prey organisms so that the Myxococcus cells can “feed” in a nutrient rich soup. Finally, when nutrient conditions become limiting, this organism aggregates into high-density clusters of 50-100,000 cells, which form mound-like fruiting bodies. Some investigators have described fruiting body formation as an altruistic process. Only about 10% of the cells in an aggregate develop into stress resistant spores. Some relatively small number of cells form the capsule of the fruiting body and virtually all the rest of the cells lyse. Some investigators maintain that the lysate released by these dying cells is important to the formation of the developing spores. It would appear that most of the cells in any aggregate relinquish their ability to reproduce and contribute to the formation of a minority population of resistant spores within the fruiting body.

The role of the fruiting body is a matter of some conjecture. Some researchers suggest that the structure may improve the chances of dispersal of the spores to a new nutrient rich location. Others suggest that the tight packing of the spores and their mutual adhesion in the fruiting body may result in the clustering of spores so that many germinate together and grow to critical mass faster when an abundant supply of nutrient becomes available.
Not all examples of altruism are related to the physical structure of cellular aggregates or biofilms. In an interesting pair of papers, Baty et al. described a partitioning in biofilms of the marine bacterium Pseudoalteromonas sp S91. This differentiation was physiological rather than structural. These investigators were looking at the growth of S91 on artificial chitin surfaces using a green fluorescent (gfp) protein reporter gene linked to the ChiA locus. This gene is necessary for the utilization of high molecular weight Chitin. The enzyme controlled by ChiA is responsible for the hydrolysis of β 1-4 N-Acetylglucosamine (Chitin). The gfp reporter turns on when the gene is expressed and is detectable under UV light as a green fluorescence localized to individual cells. Investigators found that about 57% of cells attached to the chitin surface were up regulated for ChiA after 150 hours as determined by gfp expression. About 42% of the surface attached cells were down regulated for ChiA expression. On the other hand, virtually all of the cells released into the bulk fluid exhibited no ChiA activity, suggesting an adaptation in which some cells undertake the hydrolysis of the substrate while others utilize the excess N-acetylglucosamine without expressing the ChiA gene. The “pioneer” cells that detach from the biofilm and which are responsible for the initiation of other biofilm sites downstream are drawn almost entirely from this latter population.

Baty and his associates suggested that “Such altruistic behavior of cells in a bacterial population may be an adaptive response to conserve energy in oligotrophic environments such as the ocean which, when coordinated with a process like detachment for dispersal, optimizes exposure to the widely dispersed detrital POM (Particulate Organic Material) in the pelagic (oceanic) water column.”
Cheating and Cheaters
In any form of communication between individuals be they birds, bison or bacteria, it is important that the information being conveyed be “honest”, that is accurate, from the point of view of both the individual giving and receiving the signal. Nevertheless, there are circumstances when “cheating” could bring a decided advantage to the cheater. In bacterial quorum sensing, for example, a mutant, that ignored a signal to refrain from growing into the water channels of a biofilm might gain significant short-term advantages by exploiting this common resource, gaining all the advantage of cellular reproduction while sharing only a portion of the cost in terms of nutrient accessibility. If cheating can bring advantages to the cheater, how does honest communication persist? Why doesn’t natural selection act in favor of the cheaters?
In order for quorum sensing based regulatory processes to persist in nature two sorts of rules must apply. First, the “signals [must] convey accurate information about the signaler, its future behavior, or its environment.” Then, the receiver must be able to “rely” upon the information as truly representing the “intent” of the signaler. “We expect the rules determining the honesty of a signal to be imposed by natural selection, with costs and benefits ultimately measured in terms of relative reproductive success” (Searcy and Nowicki, 2008).
But as with any sort of communication there is, at least in theory, the possibility for signalers to lie and for receivers to cheat. In biofilms, as described above, there are channels, which permit the bulk fluid access to the interior of the biofilm. This fluid carries in nutrient and carries away metabolic wastes serving much the function of a circulatory system in multicellular organisms. It is to the advantage of all community members to keep the channels open. In this context the channels are a commons” in the sense of Garrett Hardin’s famous essay The Tragedy of the Commons (1968). Certain members of the biofilm population emit signals that convey the message “keep the channels open” and this message, if obeyed, is advantageous to all community members. If, however, certain individuals can ignore the signal partially or completely, they may continue to reproduce and grow into the channels increasing their numbers at the expense of other community members who are obeying the message. Of course if the channels are overgrown completely nutrient supplies are cut off and toxic wastes accumulate to the detriment of all. This is the Tragedy of the Commons Harden wrote about. But if, the cheater can encroach on the water channels just a bit, it benefits by its increased reproductive success while it bears but a portion of the burden of decreased nutrient and waste exchange.
Examples of proposed cheating behavior are being described with increasing frequency among many diverse groups of microorganisms including the cellular slime molds (Dictyostelium) and bacteria (e.g. Pseudomonas aeruginosa, Myxococcus, and Bacillus).
Perhaps the most important clinical example of cheating is found among mutants of Pseudomonas aeruginosa isolated both in vitro and from specimens obtained from patients suffering from the hereditary disease cystic fibrosis. A child inheriting the CFTR (cystic fibrosis transmembrane conductance regulator) allele from both parents is deficient in a chloride channel conducting protein located in the membranes of normal individuals. In most patients bacterial infection occurs for the first time within a few months of birth. P. aeruginosa and Burkholderia cepacia are the dominant opportunist pathogens in most cases with Pseudomonas being responsible for morbidity and mortality in 60-80% of cases (Chernish, R. N. and S. D. Aaron, 2003).
This organism’s resistance to antibiotics, its hypermutability and its formation of biofilms with large amounts of extracellular polysaccharide are all factors that contribute to morbidity and the difficulty in managing this disease. It has been found that over the course of infection the strain of P. aeruginosa undergoes sequential mutation resulting in a variety of phenotypes among the related but genetically distinct strains. Among these derived strains investigators have found many have defects in the lasR gene which controls the synthesis of the LasR transcriptional regulator protein that normally responds to the quorum sensing autoinducer molecule (3-oxo-C12-HSL). In the absence of a functional LasR protein the mutant strain becomes signal blind and fails to respond to normal QS signaling. Initially this was interpreted as evidence that Quorum Sensing was irrelevant in Pseudomonas infections in CF patients.
With the discovery that LasR mutants gain a growth advantage in rich artificial media and within the CF airway these mutants are now being reinterpreted as Quorum Sensing “Cheaters”. This growth advantage may be responsible for the persistence of the lasR mutations. The production of the QS signaling apparatus imposes both an energetic burden and a growth restriction on wild type Pseudomonas strains. Mutants, which escape this control, benefit from the control imposed on wildtype cells while escaping the necessity of making these proteins themselves. In addition to energetic advantages, cheaters are more resistant to cell death and lysis in the stationary phase of growth and exhibit increased β-lactamase activity making them more resistant to ceftazidime a β-lactam antibiotic commonly used in chronic CF airway infections Sandoz et. al 2007 and Griffin et. al .
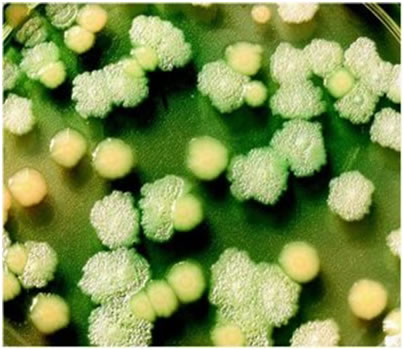
Figure 3 shows normal and mutant Pseudomonas: The rounded, yellow colonies are of a normal (wild-type) Pseudomonas colony from a cystic fibrosis patient, and the flattened, pock-marked, iridescent colonies are lasR mutant Pseudomonas colonies from the same patient. Both are growing on the same nutrient agar petri dish. Mutations that enable Pseudomonas to thrive in cystic fibrosis lung secretions may also guard the bacteria against antibiotics.
Do these tales qualify as examples of altruism and cheating? The verdict is still out, but these phenomena are certainly notable and worthy of our attention.

